
The genetics of cloaks
We talked in another article about the possible aspects of the cat's coat, with particular attention to the Maine Coon. All this variety of colors is actually governed by a few simple laws. However, it is necessary to know some notions of basis of genetics to understand how they work. The speech may seem a little complicated, but don't be discouraged!

The characteristics of every living being are established by the genetic makeup, to which the father and mother contribute equally. The genetic makeup consists of pairs of identical chromosomes, schematized as two Xs, except for the one that determines the sex. In fact, in this case, one of the two chromosomes loses a "paw", and passes from the X shape to a Y shape.
When a new living being is generated, its sex is female if the combination is XX, male if the combination is XY.
The way in which genes are combined is often visualized with the so-called Punnet square. Since it is a simple but essential tool for working with genetics, let's see how it is built in the case of sex determination. Just draw a 2x2 grid, and on the top side write the genes of the mother, and on the left side those of the father. Obviously, it can also be done vice versa.

Then in each square of each column of the grid the gene corresponding to that column is reported, in this case the pink X of the mother in the first column and again the pink X of the mother in the second. Then, in each square of the row, the gene relative to that row is transcribed, that is the blue X of the father and the blue Y of the father.
In the individual squares of the grid we then obtain the possible combinations of the father and mother genes. The combination XX indicates a female, that XY indicates a male.
In this example, it is clear that the probability of having a male or female child is 50%, but it must absolutely be emphasized that this probability begins to make sense only for a large number of events, i.e. of puppies in the same litter, approximately starting from 10 .
Since every event (birth) is independent of every other, the total statistic only begins to make sense for very large numbers of parts, say, to fix ideas, about ten . This consideration is valid for any statistical evaluation, and it is important to keep this in mind.
On each pair of chromosomes, there are sequences of tens of thousands of molecules that determine all the characteristics of the living being. Each chromosome sequence controls a single function, for example one sequence the shape of the eyes, another the length of the femur, etc. etc. These single sequences are called genes .
Pairs of chromosomes are functionally equal, meaning that each chromosome in the pair has the same sequence of genes, and each of the two genes on the pair's two chromosomes controls exactly the same function. The genes, however, are not exactly identical, since one of the pair's chromosomes derives from the mother, and one from the father, and can therefore differ in their status. In fact, genes can be in two states, which for simplicity we will call " on " or " off ". If the gene is switched on, and this is expressed by indicating the gene with a capital letter, the corresponding function is active, if it is switched off, the corresponding function is not active.
From the combination of the two genes in the chromosome pair, the overall action of the function results. For a generic gene G there can therefore be the combination GG (both genes on) Gg or gG (one on, one off), and that gg (both genes off). The on gene is referred to as the dominant gene, the off gene as recessive.
To express a characteristic linked to a dominant gene, the corresponding gene only needs to be present. In other words, if our hypothetical gene controls the presence of green eyes, all cats in which the combination GG, Gg or gG is present will have green eyes. The combination GG is called homozygous, since the genes are in the same state, that Gg or gG heterozygous, since the genes are in two different states. The two combinations Gg and gG are exactly equivalent and therefore only one is indicated. The combination gg, on the other hand, also homozygous, gives rise to a non-green color. This somewhat strange statement only says that the color of the cat's eyes will depend on the rest of the genetic make-up, but that they are certainly not green.
So, to recap, a particular characteristic linked to a dominant gene will manifest itself in 75% of cases ( always assuming a very high number of events, at the infinite limit ), that linked to the recessive gene in 25%. In the case of the dominant gene, however, only one in three subjects will be homozygous, the other two will carry the recessive gene in their genetic heritage, which can manifest itself in future generations if it is combined with another recessive gene. It therefore becomes important to distinguish between genotype , that is the set of genetic makeup, and phenotype , that is, the set of external characteristics that manifest themselves.
Having clarified the role of genes, we can now see how this affects the coat color of cats. The original coat of the cat is black brindle mackerel, therefore with each hair in alternating light and dark bands (agouti) and brindle with vertical thin lines, where areas with banded hair and areas with uniform hair alternate. The black color is expressed thanks to a dominant Black gene. In Maine Coons, the only combination allowed is the dominant heterozygous BB. In other breeds, such as the Sacred of Burma, there are also other variants, linked to the fact that B has two possible recessive genes, b and bl. The first, in the homozygous recessive combination bb, gives the color chocolate, the second blbl gives the color cinnamon (cinnamon).
In the course of evolution, a genetic mutation has caused the black pigment typical of the ancestral cat to change, under the action of a new gene, into a red pigment. This effect, managed by the mutated Orange gene, makes areas of the hair that were originally black red. In other words, in the hair there is no longer an alternation of black and light bands, but of red and light bands. This gene, when turned on (therefore O large), covers the action of the Black gene, so it is able to transform black into red even in its heterozygous Oo expression (O large or small). This covering action is technically called epistasis (from the Greek epistasis, dominant position), since one gene masks the action of another gene. In its homozygous expression oo (or small or small), the cat remains non-red, and therefore black.
Nature, however, wanted to make us a beautiful one joke, and he located this new mutated Orange gene right on the paw of the sex chromosome X which is lost when it changes to Y. So, the gene for red is carried only by the X chromosome, and is then a sex-linked gene . In this case, the Punnet square becomes a little more complicated, since the sex of the cat must also be taken into account. Let's see what happens by putting together a black XY o- male and a red XX OO female.
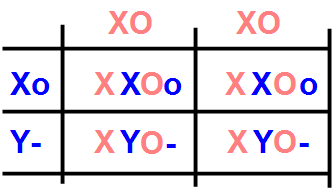
First, we see that the black cat has only one gene for the red color, in its recessive form or. The other gene in the pair is missing, as the male has only one X chromosome, and is denoted by a hyphen "-". The female, on the other hand, contributes both O chromosomes.
Let's look first at the second row of the square, where there is only the combination of the XY sex chromosomes. The males that are born from this mating are both red (O-), so they inherit the color of the mother. On the first line there are only XX combinations, therefore females, which have both O and O genes, one inherited from the mother, one from the father. What color will these Oo bitches be? In theory, the O is dominant, so they should be red. In practice, a particular phenomenon occurs during the growth phase of the embryo, linked to the fact that the Orange gene is on the sex chromosome X. In fact, while females have two X chromosomes, the male has only one, and therefore all the other genes present on the X chromosome have half the possibility of gene expression in the male. Put simply, the male's genes on the X chromosome would produce half the proteins of the female's genes, resulting in an obvious competitive disadvantage. Mother Nature has remedied this problem by ensuring that, in all cells of the female, during embryonic development one of the two X chromosomes is deactivated, absolutely randomly, cell by cell and, for all cells, at a random moment during the development of the fetus . The result, from the point of view of the coat, is that the cells responsible for the production of the pigment of the hair will be able to produce either red pigment, or black pigment, giving rise to that mixture of red and black colors typical of tortie . If inactivation occurs early, when there are still few cells, the subsequent duplications of a cell in which one of the two genes has been activated, say that for the red color, maintain the same color, and similarly for black. This produces a coat with large red and black spots. If, on the other hand, the inactivation occurs late, the two colors appear well mixed. So, in summary, from the mating between a black male and a red female, tortie females and red males are born .
The expression of black and red colors through the Black and Orange genes can be modified by the action of the Dense gene. In the most common condition, with the DD pair, the cat has black or red color. In its homozygous recessive dd version, the distribution of the pigment in the hair becomes uneven, with the result of having greater reflection of light and therefore a more diluted, less intense color. The genetic makeup BB-oo-dd gives rise to blue-gray cats, while that of BB-OO-dd to cream-colored cats, and that of BB-Oo-dd to blue-cream tortie.
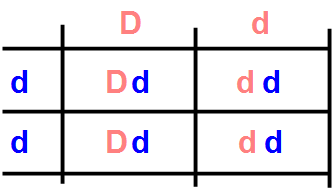
The fact that diluted color occurs only when the Dense gene is in its homozygous recessive form dd makes it necessary to mate both cats with diluted color to obtain puppies with diluted color. However, this can happen even if one of the parents does not express the dilution gene, but carries it in his genetic patrimony, the result for example of an ancestor. In fact, by mating a Dd with a dd, from the relative Punnet square we see that, statistically, half of the puppies have diluted color, and half do not, even if they carry the relative recessive gene.
Another epistatic gene, that is, that masks the action of other genes, is that linked to the presence of white spots in the coat. This gene, called piebald Spotting , masks the expression of the Orange and Black genes in those areas. This gene acts during embryonic development, preventing the cells that generate the pigment from reaching their position. If there is hair on the areas where the gene acts, this hair is white. If, on the other hand, there is skin, such as on the nose or on the fingertips, this is depigmented in pink. The gene is dominant, although its expression is variable. There are strong indications that the dominant homozygous version SS gives white areas of greater extension than the heterozygous version Ss, although the quantity and the localization of the spots of the children starting from that of the parents is currently unpredictable. The homozygous recessive version ss obviously does not give white spots.
A reinforced version of the S gene is that of the White gene, which results in a completely white coat. It is a dominant gene, so both the WW and the Ww combination give the same result. The mechanism of action is the same as the S gene, except that it affects the entire coat of the cat. However, other effects occur. In fact, the embryonic cells that originate the skin pigment also generate the pigment of the fundus of the eye and the pigment cells of the inner ear. If the action of the gene, in the phase of the initial separation of the cells, is aimed at the cells responsible for the formation of the ocular tissue, the inhibition of the pigmentation results in the fact that that eye will have, at the end of the growth, a blue or light blue color. clear. Of course, it can happen that one eye is depigmented and the other is not, resulting in the fascinating unequal eyes. If the action of the gene also affects the ear tissues, you may have malfunction or deafness, mono or bilateral. However, there are no certainties in this sense, and the inheritance rules of eye color and deafness are still to be established, and only a few statistical indications can help us.

The first thing to note is that if the puppy has colored spots on his head, a trace of the color masked by the W gene, the probability that he is deaf is around 30%, while it rises to 50% if the spots are not there. The possible explanation is that if a few hair pigment cells have arrived at their destination, it is plausible that the same has happened for those of the inner ear, sometimes enough to avoid deafness. The second correlation is that with eye color.
Based on a series of studies conducted in various countries, and shown in the left figure, cats with both yellow eyes have a 10-20% probability of deafness, cats with unequal eyes 30-40% probability, cats with both blue eyes probability of deafness around 60-80%. Note that when it comes to deafness, it can be both mono and bilateral, equivalent from a genetic point of view.
So far we have implicitly only dealt with cats with solid color, that is, without any branding. As mentioned in another article , the branding design depends on two conditions; one is the presence of agouti hair, i.e. in which the single hair has no uniform color but alternating light and dark bands, the other is the presence of alternating bands where agouti hair is present (with alternating light and dark bands and therefore more light) and non-agouti hair (uniform and therefore darker hair). The areas where the hair is darker are called markings (streaks) of the brindle. Each of these characters is controlled by a gene. Whether the hair is banded or not depends on the Agouti gene. The ancestral cat has agouti hair, and therefore the dominant expression AA is the natural one, while the recessive combination aa makes the hair uniform. So in the cat AA, or Aa (dominant agouti hair) there is the banded hair, and therefore the branding designs are possible, while the aa cat has no agouti hair and is not (or should not be) brindle. The tabby design, on the other hand, is managed by the Tabby gene. In its homozygous dominant form TT gives rise to the tabby mackerel pattern, with parallel lines. Its recessive form is indicated with tb-tb, as it gives rise to the classic tabby or blotched design. The intermediate spotted form is not due to a particular gene, but to a set of several modifier genes, not yet identified, which interrupt the parallel lines of the mackerel and originate the spotted pattern. The same modifier genes are responsible for the incomplete dominance of mackerel over the blotched. The T-tb genotype is often a poorly defined mackerel, with the two patterns mixing together. For completeness, we mention the Ticked Ta gene, which when expressed in a dominant form (TaTa or Tata) is epistatic (covers) all the other genes responsible for the design. This brindle design gives rise, for example, to the classic Abyssinian cat coat.
It must also be remembered that the Agouti gene and the Tabby gene are different genes, and that therefore a solid cat carries with it, at the genotype level, the tabby gene, whatever it is, and that this can be expressed again if the progeny were brindle. Furthermore, in solid black puppies with an unexpressed Tabby gene, a ghost brindle is still recognizable, which is often still identifiable as an adult, looking at the hair with grazing light. In cats with the dominant Orange gene, the action of the non-agouti aa gene is absolutely ineffective, so much so that it is extremely difficult to have solid red cats. This is also evident in the brindle tortie, which have much clearer and more evident branding in the black areas than in the red ones.
As a last topic, let's face the silver colors, where there is a whitening of the base of the hair, which can go from one third to two thirds of the length of the hair itself. In the first case, in cats with solid color, we speak of smoke color, in the second of silver. In brindle cats, the silver effect is to lighten the base of the hair in the marking of the stripes, and to make the bands of the hair silver in the areas with agouti hair between the markings. In Maine Coons, the single gene theory accounts for almost all phenotypic aspects of the coat and so we will rely on that. The responsible gene is called the melanin inhibitor gene, and in its dominant expression II or Ii it generates the whitening of the hair at the base, while in the recessive version ii it gives no effect. Based on this simple model, it is possible to predict the presence of silver in offspring, but not account for all the details, such as contrast and extent of bleaching. The genetic factors involved in producing the ideal silver, solid or not, are complex. Not only are there many interacting genes, but genes sometimes don't express themselves fully, or conflict with each other. For example, the melanin inhibitor gene sometimes does a bad job of blocking pigment production, resulting in an undercoat that is too gray or yellowish in color. Similarly, poorly expressed non-agouti or overexpression of the melanin inhibitor gene can generate a pale, washed-out smoke coat. Various polygens (sets of related genes), factors related to environmental influence on gene expression (epigenesis), or modifier genes not yet identified are believed to lead to different staining phenotypes, some deemed more desirable than others by humans. breeders.